Reza Ranjbar1,
Samanehsadat Sharifimoghadam3,
Elnaz Saeidi3,
Nematollah Jonaidi2,
Amirhossein Sheikhshahrokh3 ![]()
For correspondence:- Amirhossein Sheikhshahrokh Email: Amir.genetics@yahoo.com Tel:+989365819491
Received: 31 May 2016 Accepted: 11 September 2016 Published: 31 October 2016
Citation: Ranjbar R, Sharifimoghadam S, Saeidi E, Jonaidi N, Sheikhshahrokh A. Induction of protective immune responses in mice by double DNA vaccine encoding of Brucella melitensis Omp31 and Escherichia coli Eae genes. Trop J Pharm Res 2016; 15(10):2077-2083 doi: 10.4314/tjpr.v15i10.4
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the efficacy of a double DNA vaccine encoding of Brucella melitensis omp31 gene and of Escherichia coli eae gene in inducing protective immune response in a mouse model.
Methods: After performing PCR assays and cloning both the eae and omp31 genes, the generated DNA vaccines were intramuscularly injected into BALB/c mice. Purified proteins and transformed cell lysates were identified by western blot and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and analyzed by enzyme-linked immunosorbent assay (ELISA).
Results: pcDNA3-omp31-eae provokes a significant, high level of protection in BALB/c mice against challenges with E. coli and B. melitensis. Titers ranging from 90:160 to 2,570:5,110 were detected 60 days after vaccination. Notably, rOmp31 and rEae proteins significantly induced the production of IFN-γ in cells from pcDNA3-omp31-eae-immunized mice (p < 0.05 in both groups) and only low levels of IL-10 were found in all groups. The lymphocytes from mice immunized with pcDNA3-omp31-eae had an apparent T cell proliferative response to rOmp31 and rEae proteins (p < 0.01) with a stimulation index of 14.90.
Conclusion: The novel pcDNA3-omp31-eae construct can be used as a DNA vaccine to prevent E. coli and B. melitensis infections in future studies.
Introduction
DNA vaccines can induce both cellular and humoral immune responses, but have demonstrated only limited success in animals because of their low immunogenicity [1]. Nonetheless, DNA vaccines have several advantages over traditional protein-based vaccines, including induction of longer lasting immunity, ease of development, and lower production costs [2].
One of the most widespread zoonoses, caused by the pathogenic bacterium Brucella, is brucellosis, a disease that affects humans and livestock. Brucella melitensis is an intracellular, facultative, Gram-negative bacterium that infects humans and a wide range of animals [3,4]. B. melitensis has high pathogenicity in humans and causes a large proportion of human cases [5,6]. Human brucellosis is treatable with antibiotics, but the course of antibiotic treatment must be long because Brucella infections are intracellular [7]. Brucella species are therefore considered potential civilian, military, and agricultural bioterrorism agents [7]. In livestock, brucellosis can cause great economic losses, as infection can lead to stillbirths, abortions, and loss of fertility in animals [8,9].
B. melitensis causes miscarriage in cows, goats, and sheep, and is considered to be the most pathogenic Brucella sp. for humans [10]. Recombinant Omp31 (rOmp31) vaccination elicits a CD4 Th1 response that provides partial protection against B. melitensis infections [11]. High concentrations of the plasmid containing the omp31 gene are needed to generate an efficient response, probably because of the low in vivo transfection efficiency for this type of plasmid vector [3].
Infections with Escherichia coli are commonly associated with diarrhea accompanied by bloody hemorrhagic colitis and abdominal cramps, but more serious complications such as thrombotic thrombocytopenic purpura, hemolytic-uremic syndrome, hemorrhagic colitis (HC), and in some cases, death, can also occur [12-14]. Pathogenic strains of E. coli are classified as either extra-intestinal (ExPEC), causing a variety of infections in both animals and humans, or intestinal pathogens causing diarrhea [15]. The chromosomal gene eae, which encodes intimin, is part of a pathogenicity island termed the locus of enterocyte effacement (LEE) [13]. Eae is the first gene to be associated with attaching and effacing (AE) activity, and is essential for intimate bacterial attachment to eukaryotic host cells [14]. The eae gene is one of the genes currently used for the molecular diagnosis of enteropathogenic E. coli (EPEC) [16]. EPEC produces characteristic AE lesions on intestinal epithelial cells. The AE phenotype is encoded by a cluster of genes, including the eae gene located on the LEE [17]. The ability to cause AE lesions in enterocytes is the major virulence factor of E. coli in intestinal infections in humans and animals [18].
The E. coli AE gene (eae) encodes a 94-kDa outer membrane protein, intimin, which is important but not sufficient for the production of AE lesions [18,19]. Inoculation with plasmid DNAs expressing foreign proteins stimulates both cellular and humoral immune responses that protect against bacteria, parasites, viruses, and tumors [20], and is a reasonably powerful and novel technique of immunization [21]. The aim of the present study is to investigate the immunogenicity and protective capacity of a DNA vaccine encoding the eae and omp31 proteins of E. coli and B. melitensis, respectively.
Methods
Bacterial strains and DNA extraction
B. melitensis and E. coli were obtained from the Microbiology Research Center of Islamic Azad University, Shahrekord Branch, Shahrekord, Iran. The B. melitensis and E. coli strains were grown on Brucella agar and Luria-Bertani (LB) agar, respectively. Genomic DNA was extracted using a Qiagen DNA extraction kit (Qiagen, Ltd., Crawley, UK). Quality was assessed according to manufacturer’s instructions by electrophoresing the extracted DNA on a 1 % agarose gel and measuring the optical density at 260 nm (OD 260) d [22,23].
PCR amplification
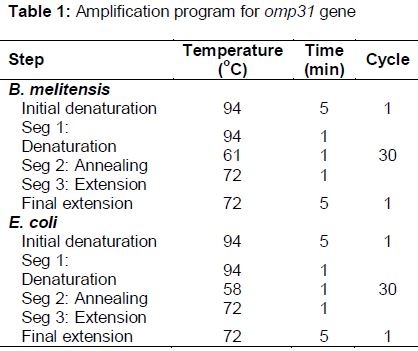
Specific oligonucleotide primers for the omp-31 gene (omp31-F: 5'-ACCGGATCCACCACCATGA AATCCGTAATTTTG-3' and omp31-R: 5'-TGCTC GAGGAACTTGTAGTTCAGACCGAC-3') and the eae gene (eae-F: 5'-ACACTCGAGCAAACCAA GGCCAGCATTAC-3' and eae-R: 5'-CAGTCTA GATTATTCTACACAAACCGCAT-3') were designed and used in their respective amplification reactions. To amplify the omp31 and eae genes, the 25-μL total volume PCR mixtures consisted of 2.5 µL 10× PCR buffer, 1 μM each of the appropriate primers, 2 mM MgCl2, 200 μM dNTPs, 1 μg template cDNA, and 1 unit pfu DNA polymerase (Roche Applied Science, Penzberg, Germany). For each gene, the 30-cycle amplification was performed in a thermal cycler system (Palm Cycler Gradient, Sydney, Australia) with their respective amplification programs are shown in .
Cloning of bacterial genes
Electrophoresis was subsequently performed on a 1 % agarose gel in 1× TBE buffer, followed by staining with ethidium bromide and visualization on a UV transilluminator to verify the PCR product sizes. Bands corresponding to the omp31 and eae genes were cut from the gel and the DNA fragments were purified using a gel extraction kit (Bioneer, Cheongwon, Korea) according to the manufacturer’s instructions. TOPO-omp31 and TOPO-eae plasmids were cloned using the TA technique, as follows. Omp31 and eae DNA fragments were ligated to the T-vector using the TOPO T/A cloning kit (pCR8/GW/TOPO, Invitrogen, Carlsbad, California, United States) according to the manufacturer’s instructions, to obtain TOPO-omp31 and TOPO-eae. BamHI/XhoI and XhoI/XbaI restriction analysis and PCR techniques were used to confirm insertion. The recombinant plasmids (TOPO-omp31 and TOPO-eae) were sequenced with specific primers using the Sanger method (Macrogen, Seoul, Korea), then linearized using BamHI/XhoI. The omp31 and eae genes were released from TOPO-omp31 and TOPO-eae, respectively, using BamHI/XhoI and XhoI/XbaI and inserted into the pcDNA3 plasmid. The pcDNA3-omp31-eae plasmid constructs were verified by restriction digestion and sequencing.
Preparation of recombinant omp31 and eae proteins
To prepare recombinant Omp31 (rOmp31) and Eae (rEae) proteins, the omp31 and eae genes were excised from the pcDNA3-omp31-eae plasmid by BamHI/XhoI and XhoI/XbaI digestion and then inserted into the same restriction sites of the prokaryotic expression vector pET32a(+) (Novagen, Wisconsin, United States). The recombinant plasmids were transformed into E. coli BL21 (DE3) (Novagen). After culturing, the positive clones were selected. According to the manufacturer’s protocol, the recombinant proteins were expressed in successfully transformed bacteria by induction with isopropyl-β-D-thiogalactopyranoside (IPTG) in LB medium and purified on a Ni2+-HiTrap chelating 5-mL pre-packed column (Amersham Pharmacia Biotech, Piscataway, United States) using imidazole as the elution reagent.
SDS-PAGE analysis
The purified proteins and transformed cell lysates were identified by western blot and sodium dodecyl sulfate-polyacrylamide gel electropho-resis (SDS-PAGE). Purified proteins were stored at –70 °C until use for in vitro stimulation of splenocytes or for enzyme-linked immunosorbent assays (ELISAs).
In vivo analysis
Mice were anesthetized by inhalation of 3 % halothane in 1 L/min fresh gas flow composed of an equal N2O:O2 mixture and maintenance in 1–1.5 % halothane. The tibialis anterior muscle was then injected with 50 μg of plasmid DNA (pcDNA3-omp31-eae, 100 μg DNA/mouse) in 50 μL of phosphate buffered saline (PBS) using a 28-gauge needle insulin syringe. Mice were vaccinated at weeks 0, 2, and 4 with pcDNA3-omp31-eae. As negative controls, separate groups of mice were immunized with pcDNA3, with PBS only, or with plasmid lacking either the omp31 or eae inserts. As a positive control, another group of mice was vaccinated intraperitoneally with 2 × 108 CFU of B. melitensis strain Rev 1 and E. coli in 0.2 mL PBS.
Analysis of IgG isotypes
The presence of serum immunoglobulin G (IgG) isotypes specific for rOmp31 and rEae was determined by indirect ELISA. pPurified rOmp31 and rEae (3 μg mL-1 in carbonate buffer, pH 9.6) were added to standard polystyrene 96-well plates for ELISA (Nunc-Immuno plates with MaxiSorp, Nunk, Roskilde, Denmark), incubated overnight at 4 °C, then washed four times in wash buffer (Tris-buffered saline, pH 7.4, with 0.05 % Tween 20). The plates were blocked with 5 % gelatin in Tris-buffered saline at 37 °C for 2 h, then incubated for 3 h at 24 °C and washed four times with serially diluted sera. Isotype-specific rabbit anti-mouse horseradish peroxidase conjugates were added (ICN Biomedicals, Inc., Aurora, OH, USA) (50 μL well-1) at appropriate dilutions. Plates were washed four times and 100 μL of substrate solution (200 μmol of o-phenylenediamine and 0.04 % H2O2) was added to each well after 30 min, followed by incubation at 24 °C. The enzyme reaction was stopped after 20 min incubation at room temperature by the addition of 100 μL 0.18 M sulfuric acid, and the absorbance at 492 nm was measured.
Mice were sacrificed two weeks after the last immunization and spleens were removed under aseptic conditions. After preparation of single-cell suspensions from the spleens, red blood cells were lysed with ACK solution (150 mM NH4Cl, 1 mM KHCO3, 0.1 mM Na2·EDTA, pH 7.3). Splenocytes were cultured in 5 % CO2 at 37 °C at a concentration of 4 × 105 cells well-1 in RPMI 1640 medium supplemented with 2 mM L-glutamine and 10 % heat-inactivated fetal calf serum (Sigma, St. Louis, United States) in a 96-well flat-bottom plate in the presence of 0.08 μg purified rOmp31 and rEae protein or no additives (unstimulated control). Cells were cultured for 3 days and pulsed for 8 h with 0.4 μCi of 3H-thymidine (50 Ci mmol-1; Amersham Pharmacia Biotech) per well. Incorporated radioactivity was measured in a liquid scintillation counter.
Cytokine detection and vaccine evaluation
For cytokine detection, spleen cell culture supernatants were tested for the presence of cytokines by antigen-capture ELISA using OptEIA Set Mouse IFN-γ, IL-4, and IL-10 (BD Biosciences, San Diego, CA, USA) and collected after 48 h of antigen stimulation.
Five weeks after vaccination, six mice from each group were challenged by intraperitoneal injection of 104 CFU of B. melitensis 16M or E. coli. After two weeks, the infected mice were sacrificed. Dilutions of spleen homogenate were plated to determine the number of E. coli and B. melitensis CFUs per spleen. This assay was performed in triplicate. Log10 units of protection were obtained by subtracting the mean log10 CFU for the experimental group from the mean log10 CFU of the corresponding control group.
Ethical consideration
This work was approved by the ethical committees of the biotechnology research center, Islamic Azad University, Shahrekord, Iran and Research Adjutancy of the Islamic Azad University of Shahrekord (ref no. 2037550IAU). The laboratory animal studies followed international ethical and scientific guidelines on animal testing and research [24].
Results
Amplified eae and omp31 genes
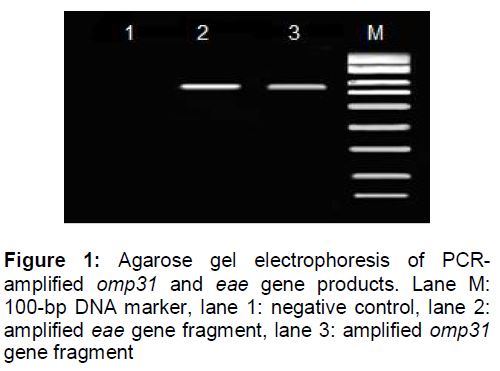
The PCR amplified products for the B. melitensis omp31 and E. coli eae genes were respectively 720 bp and 862 bp in length, as shown in .
B. melitensis omp31 and E. coli eae genes were successfully cloned into TOPO vector. In addition, positive control colonies were obtained; matrix analyses of the cultured colonies for the omp31 and eae genes are shown in Figures 2 and 3, respectively.
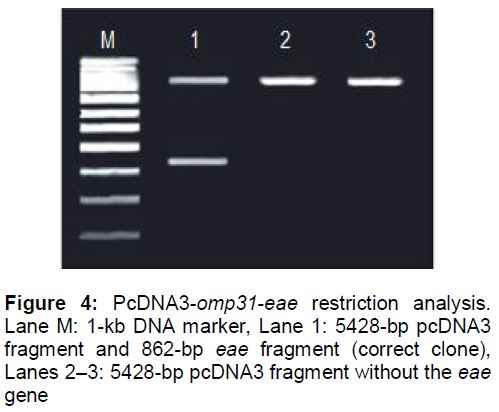
Omp31 gene fragment, which has a BamHI/XhoI restriction site, and the eae gene fragment, which has a XhoI/XbaI restriction site, were inserted in the polyclonal site (PCS) of pCDNA3. The result of pcDNA3-omp31-eae restriction using XhoI/XbaI enzymes is shown in .
Antibodies and immunological responses
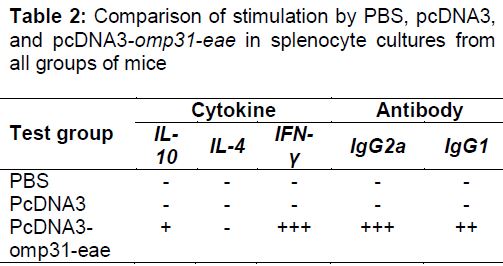
Immunization with pcDNA3-omp31-eae led to a rapid IgG response within two weeks after vaccination. Titers ranged from 90:160 to 2,570:5,110; the peak titers were detected 60 days after vaccination. IgG1 and IgG2a antibody analyses revealed that anti-Omp31 and anti-Eae antibodies were found in pcDNA3-omp31-eae immunized mice, but not those given pcDNA3. We determined the proliferation responses and cytokine profiles of spleen cells from mice injected with pcDNA3-omp31-eae, pcDNA3, and PBS to examine cell-mediated immunity (CMI). Lymphocytes from mice immunized with pcDNA3-omp31-eae had an apparent T cell proliferative response to rOmp31 and rEae proteins (p < 0.01) with a stimulation index of 14.90.
Cytokine profiles using supernatants of spleen cell cultures revealed higher levels of IFN-γ in pcDNA3-omp31-eae-immunized animals compared to the negative control groups. The rOmp31 and rEae proteins significantly induced the production of IFN-γ in cells from pcDNA3-omp31-eae-immunized mice (p < 0.05 in both groups), and only low levels of IL-10 were found in all groups. In addition, no IL-4 was detected in culture supernatants of splenocytes that were stimulated with specific antigens.
Protection tests were accomplished by challenging control and vaccinated mice with B. melitensis and E. coli and evaluating infection levels by determining CFU counts in the spleen. The results shown in indicate that immunization with pcDNA3-omp31-eae resulted in a significantly higher degree of protection in comparison with the unimmunized control groups.
Discussion
DNA vaccines consisting of genes cloned into a eukaryotic plasmid are a novel and promising approach that can induce both humoral and cellular immune responses [25]. Control of Escherichia and Brucella has markedly reduced the incidence of human infection, but these diseases still represent an important cause of morbidity worldwide. To design a new generation of vaccines, more information on the antigenic makeup of Brucella and Escherichia spp. must be obtained to identify immunodominant proteins and epitopes. A human vaccine would be valuable for individuals who may be occupationally exposed to Brucella and Escherichia spp.
Numerous attempts have been made to control human disease by vaccination with attenuated Brucella strains [21]. However, practical acceptance of these preparations has been very limited, mainly because of the unacceptable local inflammatory responses they elicit, combined with their limited protection. Obviously, there is a need for a better vaccine for brucellosis eradication [21]. Mice immunized with omp31 were significantly protected against B. ovis and B. melitensis infections [3].
Eae gene cloning results of the present study were similarly successful as those of for expression of the STX2 gene of E. coli O157:H7 in pUC118 and pUC119 vectors [12] and of Hongyan et al [26]. Thus, our DNA construct can be used as a DNA vaccine against E. coli in future studies.
The development of a safe and effective vaccine against B. melitensis and E. coli has proven to be complicated. Indeed, numerous intracellular and cell surface components have been assessed as protective antigens against these two bacterial infections, although only low or intermediate levels of protection have been achieved with these isolated proteins [9,12]. The B. melitensis omp31 and E. coli eae genes are promising candidates for use as a subunit vaccine against brucellosis and E. coli infection. In the case of B. melitensis and E. coli, cellular immune responses characterized by the production of IFN-γ are critical to conferring protection [27].
In summary, this study demonstrates that DNA vaccines can be effective against E. coli and B. melitensis infections. In this study, we also assessed the capacity of pcDNA3-omp31-eae to elicit an immune response and protective immunity in BALB/c mice. Since the levels of protection achieved by a multiple DNA vaccine encoding the omp31 gene of B. melitensis and the eae gene of E. coli are more than sufficient, this double DNA vaccine could be a useful candidate for the development of DNA vaccines against E. coli and B. melitensis infections.
Limitations of the study
DNA vaccines are limited to protein immunogens, that is, they can’t be used to confer immunity to non-protein based antigens such as bacterial polysaccharide. Induced antibody production against DNA may occur in the host organism. The achieved immune response in mice was satisfactory, but to obtain immunization in larger animals, high-level DNA doses are needed. In addition, insertion of foreign DNA into the host genome in order to obtain immune response may affect the control of cell growth.
Conclusion
The DNA construct that was produced in this work can be further developed as a DNA vaccine against E. coli and B. melitensis. In addition, further studies are required to determine the functions of various T cell types in the protection induced by pcDNA3-omp31-eae.
Declarations
Acknowledgement
References
Archives


News Updates